Complesso di Golgi: terminologia, occorrenza, morfologia, struttura e altri dettagli
Complesso di Golgi: terminologia, occorrenza, morfologia, struttura, polarizzazione strutturale e biochimica, composizione chimica, funzioni e origini!
Nel 1898 per mezzo di un metodo di colorazione dell'argento, Golgi scoprì una struttura reticolare nel citoplasma. Il nome "apparato di Golgi" generalmente dato a questa struttura è confuso perché suggerisce una relazione definita con i processi fisiologici della cellula.
Oggi sembra più appropriato usare il nome "sostanza di Golgi" o "complesso di Golgi" per riferirsi a questo materiale che ha proprietà di colorazione speciali. Poiché il suo indice di rifrazione è simile a quello della matrice, il complesso di Golgi è difficile da osservare nelle cellule viventi. L'uso del microscopio elettronico ha fornito un'immagine distinta di questo componente e la sua struttura sub-microscopica è stata rivelata.
Per anni si è pensato che il complesso di Golgi fosse un artefatto di varie procedure di fissazione e colorazione. In altre parole, molti scienziati ritenevano che la struttura osservata durante numerose procedure di microscopia e definita il Golgi non esistesse realmente nella cellula vivente. Guilliermond (1923), Parat (1927), Walker e A lien (1921), sorsero dubbi riguardo all'esistenza del complesso di Golgi.
Terminologia del complesso di Golgi:
Holmgren si riferiva al complesso di Golgi come trophospongium (Cajal chiamava i canali Golgi-Holmgren). Baker usava il termine lipocondrio a causa del presunto contenuto lipidico. Il termine Dalton Complex fu dato dopo il nome del suo osservatore Dalton nel 1952. Sjostrand propose il termine cito-membrane per il sistema di Golgi.
Sosa ha suggerito la seguente nomenclatura per il complesso di Golgi:
1. Golgiokinesis :
Divisione dell'apparato di Golgi durante la divisione nucleare.
2. Golgiosomes :
I corpuscoli prodotti dalla Golgiogenesi sono chiamati Golgiosomes che sono descritti come materiale di Golgi negli invertebrati.
3. Golgiolisi :
Processo di dissoluzione dell'apparato di Golgi.
4. Gogiorrhexis :
Frammentazione sull'apparato di Golgi.
5. Golegiogenesi :
Formazione e differenziazione del corpo del Golgi durante lo sviluppo embrionale.
6. Golgio-citoarchitettura :
Studio della struttura della cellula in relazione all'apparato di Golgi.
Evento:
Il complesso del Golgi si verifica in tutte le cellule eccetto le cellule procariotiche (cioè, micoplasmi, batteri e alghe blu verdi) e le cellule eucariotiche di alcuni funghi, spermatozoi di briofite e pteridofite, cellule di provette mature di piante e sperma maturo e sangue rosso cellule di animali.
Morfologia del complesso di Golgi:
La morfologia del complesso di Golgi varia da una cellula all'altra a seconda del tipo di cellula in cui si trovano. Sono state osservate due forme di complesso di Golgi.

1. Forma localizzata :
Nelle cellule polarizzate dei vertebrati (che hanno base e apice), il complesso di Golgi si presenta singolarmente e occupa una posizione fissa. Si trova tra il nucleo e il polo secretorio. Questo può essere visto meglio nelle cellule tiroidee, nelle cellule esocrine del pancreas e nelle mucose dell'intestino.
2. Forma diffusa :
In alcune cellule specializzate di vertebrati (cellule nervose e cellule epatiche), nella maggior parte delle cellule vegetali e nelle cellule degli invertebrati si trovano diverse unità del complesso di Golgi disseminate insieme agli elementi del reticolo endoplasmatico. Ogni unità è chiamata dictyosome. Nelle cellule epatiche si verificano fino a 50 dictosomi per cellula e in alcune cellule vegetali il loro numero può arrivare fino a centinaia.
Forma :
La forma del complesso di Golgi è piuttosto variabile in diversi tipi di animali somatici. Anche nella stessa cella ci sono variazioni in diverse fasi funzionali. La forma è, tuttavia, costante con ogni tipo di cella. Varia nella forma da una massa compatta a una rete filamentosa dispersa.
Numero:
Il numero di stacks di Golgi per cella varia enormemente, a seconda del tipo di cellula, da un minimo di uno a centinaia. C'è un solo grande in alcune celle mentre nel caso di Paramoeba ce ne sono due. In Stereomyxa (una specie di ameba) ci sono molti complessi di Golgi. Le cellule nervose, le cellule epatiche e la maggior parte delle cellule vegetali hanno anche complessi di Golgi multipli, con circa 50 cellule epatiche.
Nei rizoidi algali si vedono più di 25.000 complessi di Golgi (Sievers 1965). Il complesso del Golgi può anche rappresentare una grande frazione del volume cellulare in alcune cellule specializzate. Un esempio è la cellula calice dell'epitelio intestinale, che secerne il muco nell'intestino; le glicoproteine nel muco sono glicosilate principalmente nel complesso del Golgi.
Taglia:
Anche la dimensione è variabile. È grande nelle cellule dei nervi e delle ghiandole e piccolo nelle cellule muscolari. In generale il complesso del Golgi è ben sviluppato mentre la cellula è in stato attivo. Quando la cellula invecchia, il complesso diminuisce progressivamente di dimensioni e scompare.
Posizione:
La posizione del complesso di Golgi è relativamente fissa per ogni tipo di cellula. Nelle cellule di origine ectodermica, il complesso di Golgi è polarizzato dal tempo dello stato embrionale tra il nucleo e la periferia (Cajal, 1914). Nelle cellule esocrine secretorie che hanno in generale una tipica polarizzazione il complesso del Golgi si trova tra il nucleo e il polo secretivo.
Nelle ghiandole endocrine la polarità di questo organoide è variabile, tranne che nella tiroide, dove è orientata verso il centro del follicolo. Nelle cellule più giovani e spesso nelle più vecchie si trova più comunemente su un lato del nucleo, ma in alcuni casi può circondarlo completamente. Nelle cellule gangliari del topo la posizione è perinucleare.
Struttura dettagliata del complesso di Golgi:
Dalton e Felix (1954) descrissero il complesso di Golgi nell'epididimo del ratto dopo aver preso le prime micrografie elettroniche.
La seguente descrizione del complesso di Golgi è composita basata sul lavoro di diversi autori:
1. Cisternae :
Le cisterne o sacculi sono simili alla superficie liscia ER, e appaiono in sezione come pile di sacche delimitate da una membrana molto ravvicinata. Il numero di sacculi varia da 4 a 8 nella maggior parte dei tipi di cellule animali e vegetali. In Euglena, il numero può arrivare fino a 20.
La membrana delle saccole ha uno spessore di circa 60-70 ° in grado di racchiudere una cavità larga circa 150 °, i cui bordi sono spesso dilatati. Secondo la maggior parte degli autori, ci sono due facce ben definite delle cisterne, cioè convesse e concave; quest'ultima viene generalmente indicata come la faccia matura o formante o distale e il lato convesso è assunto come faccia immatura o eccitatrice o prossimale, le menzioni cisterane in serie parallele sono separate l'una dall'altra da uno spazio di circa 200 a 300 A 0 .
Ciò che li tiene insieme non è ancora noto, ma in poche cellule un sottile strato di elettroni opachi, materiale a volte denso è visto tra i sacculi che in certe regioni sono più prominenti a cui Amos e Grimstone (1968) hanno applicato i termini nodi. Mollenhauer et al. (1973) hanno esplorato in dettaglio alcuni elementi e placche intercostali in alcuni complessi di Golgi.

2. Tubuli :
Dall'area periferica delle cisterne si forma una rete piatta complessa e anastomatica di tubuli con diametro da 300 a 500 A °. Clowes e Juniper (1969) hanno confrontato questa rete tubolare con un disco di pizzo.
3. Vescicole :
Le vescicole sono piccole sacche simili a goccioline che rimangono attaccate ai tubuli alla periferia delle cisterne.
Sono di seguito due tipi:
(a) Vescicole lisce :
Le vescicole lisce hanno un diametro compreso tra 20 e 80 μ. Contengono materiale secretorio (così spesso vengono chiamate vescicole secretorie) e sono nascoste dalle estremità dei tubuli cisternali all'interno della rete. Spesso più di una connessione tubulare, e presumibilmente riempire, una singola vescicola formante.
(b) vescicole rivestite :
Le vescicole rivestite sono protuberanze sferiche, di circa 50 μm di diametro e con una superficie ruvida. Si trovano alla periferia dell'organello, generalmente alle estremità dei singoli tubuli e sono morfologicamente abbastanza distinti dalle vescicole secretorie. La loro funzione è sconosciuta.
4. Vacuoli di Golgian :
Queste sono grandi sacche arrotondate presenti sulla faccia in scadenza di Golgi. Questi sono formati dalle cisterne espanse o dalla fusione di vescicole secretorie. I vacuoli sono riempiti con una sostanza amorfa o granulare.
Il complesso del Golgi è strutturalmente e biochimicamente polarizzato:
Il complesso del Golgi ha due facce distinte: una faccia cis, o faccia formante e una faccia trans o maturing. La faccia cis è strettamente associata a una parte transizionale liscia dell'ER grezzo. Nelle cellule secretorie, la faccia trans è la faccia più vicina alla membrana plasmatica: qui, le grandi vescicole secretorie si trovano esclusivamente in associazione con la faccia trans di una pila di Golgi, e la membrana di una vescicola secretoria formante è spesso continua con quella di la faccia trans dell'ultima ("trans-most") cisterna.
Al contrario, le piccole vescicole di Golgi sono localizzate più uniformemente lungo lo stack, si pensa comunemente che le proteine entrino in una pila di Golgi dal pronto soccorso sul lato cis e escano per più destinazioni sul lato trans; tuttavia, né il loro percorso esatto attraverso il complesso di Golgi né il modo in cui viaggiano da cisterna a cisterna lungo ogni pila sono noti.
Le due facce del complesso di Golgi sono biochimicamente distinte. Ad esempio, una variazione nello spessore delle membrane di Golgi può essere rilevata attraverso lo stack in alcuni casi, con quelli sul lato cis più sottili (simili a ER) e quelli sul lato trans più spessi (come quelli della membrana plasmatica).
Più sorprendenti sono i risultati ottenuti quando alcuni test istochimici vengono utilizzati in congiunzione con la microscopia elettronica per localizzare particolari proteine all'interno del complesso di Golgi. Alcuni di questi test rivelano l'enzima legato alla membrana, attività che mostrano una polarità distinta nella loro localizzazione all'interno della pila di Golgi.
Una scoperta biochimica particolarmente intrigante è stata la scoperta che gli enzimi lisosomiali, come la fosfatasi acida, sono concentrati con la cisterna trans-più della pila di Golgi e all'interno di alcune delle vescicole rivestite nelle vicinanze. Questo suggerisce che le vescicole specifiche in partenza per i lisosomi sono assemblate in questa regione.
Le proteine secretorie sono trovate con metodi istochimici in tutte le cisterne impilate, anche se le grandi vescicole secretorie in cui questi prodotti sono concentrati e associati solo al cisterna del Golgi trans.
Composizione chimica:
Per quanto riguarda la composizione chimica del complesso di Golgi, è stato dimostrato che sono presenti le seguenti sostanze:
1. Fosfolipidi :
La composizione dei fosfolipidi delle membrane di Golgi è intermedia tra quelle delle membrane endoplasmatiche e le membrane plasmatiche.
2. Proteine ed enzimi :
Il complesso di Golgi di cellule vegetali e animali diverse mostra grandi variazioni nel contenuto di proteine e di enzimi. Alcuni degli enzimi sono ADPasi, ATPasi, citocromo-reduttasi NADPH, transferasi glicosica, galattosina1 transferasi, pirofosfato di tiamina ecc.
3. Carboidrati :
Entrambe le cellule vegetali e animali hanno alcuni componenti carboidrati comuni, come glucosarine, galattosio, glucosio, mannosio e fruttosio. I Golgi vegetali sono privi di acido sialico, ma si trova in concentrazioni elevate nel fegato di ratto. Alcuni carboidrati come xilulosio e arabinosio sono presenti solo nelle cellule vegetali.
4. Vitamina С :
La frazione di vitamina С immagazzinata nel complesso di Golgi è stata mostrata da Tomitte. Secondo lui il complesso di Golgi immagazzina la vitamina С e la libera lentamente nel citoplasma in quantità sufficiente a prevenire l'ossidazione dei prodotti cellulari.

Funzioni del complesso di Golgi:
1. Formazione di Acrosoma durante la Spermiogenesi :
Durante la maturazione degli spermatozoi, il complesso del Golgi gioca un ruolo nella formazione dell'acrosoma (Burgos e Fawcett, 1955).

Nelle fasi iniziali, il Golgi appare come un corpo sferico, comprendente cisterne disposte in pile parallele e numerose piccole vescicole. Il più tardi sempre rimosso dalle cisterne. Con il progredire dello sviluppo, il complesso del Golgi diventa di forma irregolare e i grandi vaculoli sono formati da dilatazioni delle sacche cisterne.
Al centro di questi grandi vacuoli o vacuoli è presente un denso granulo, il granulo proacrosomale. Questo granulo derivato dal complesso di Golgi continua a crescere all'interno del vacuolo con un processo noto come accrescimento. Questo vacuolo e granello si avvicinano al polo anteriore della membrana nucleare, costituendo un granulo acrosomiale.
Con l'allungamento dello spermatide, la vescicola acrosomiale si diffonde sulla superficie nucleare e infine collassa con la membrana nucleare, formando il materiale del cappuccio. Il granulo acrosomiale diventa l'acrosoma che si trova all'apice del nucleo e apparentemente comprende alcuni enzimi coinvolti nel processo di fecondazione.
2. Sintesi e secrezione di polisaccaridi:
Studi su cellule caliciformi mediante autoradiografia e microscopia elettronica hanno stabilito l'interrelazione tra sintesi proteica, aggiunta di carboidrati e solfatazione. Le cellule caliciformi del colon producono mucigen. Questo materiale secretorio contiene una grande percentuale di carboidrati.
Il complesso di Golgi si trova appena sopra il nucleo. Verso la superficie libera della cellula si ingrandiscono gradualmente i granuli di mucene. Le cisterne prossimali del complesso del Golgi non mostrano alcun gonfiore, ma a una certa distanza attraverso la pila le cisterne distali vengono improvvisamente convertite in granuli di mucene. Le cisterne distali si convertono continuamente in granuli di mucigen ogni 2-4 minuti. Nuove cisterne prossimali si formano in compensazione.
3. Ruolo nella secrezione:
Il complesso del Golgi è considerato svolgere un ruolo nella funzione secretoria di una cellula. Ma la domanda è questa: essi stanno secernendo o sintetizzando alcune sostanze stesse o sono semplicemente un magazzino in cui i prodotti secretori che sono secreti da qualche altra parte nella cella, sono semplicemente immagazzinati e concentrati.


Dagli studi di Palade et al. 1962 questo ciclo secretorio è ora ben definito e comprende quattro fasi in caso di cellule acinose pancreatiche e sono:
(i) Incorporazione di aminoacidi in proteine sulla superficie del reticolo endoplasmatico ruvido.
(ii) Trasferimento di queste nascenti proteine secretorie nelle cisterne del reticolo endoplasmatico ruvido.
(iii) Il trasporto intracellulare di queste proteine nel complesso di Golgi.
(iv) La migrazione dei granuli di zimogeno verso l'apice della cellula dove sono scaricati in lumen.
4. Ruolo del corpo di Golgi nell'oogenesi :
Srivastava (1965) ha dato una breve recensione sul complesso di Golgi durante l'oogenesi. Secondo Afzelius (1956), il complesso di Golgi di un uovo di riccio di mare, come visto al microscopio elettronico, è costituito da pile di lamelle che formano pareti di tasche piatte, che a volte possono essere gonfie.
Ci sono alcune indicazioni delle divisioni trasversali di questi corpi. Sotelo (1959) e Sotelo e Porter (1959) hanno descritto il complesso di Golgi nel ratto-uovo visto al microscopio elettronico e hanno trovato la localizzazione juxtra nucleare di questo organello nei primi ovociti.
Nella fase successiva, questi si risolvono in frammenti e nella terza fase, questi si muovono verso la corteccia. In tutti questi casi, la loro struttura deve essere costituita da matrici molto affollate di sottili profili doppi (sacche appiattite) e vescicole sferiche.
Nei primi ovociti il complesso è organizzato in modo compatto. Negli stadi successivi, si trovano frammenti discreti di profili, circondati da piccole vescicole, sparpagliati nella zona corticale. Nei primi ovociti, il complesso di Golgi e il centrosoma sono strettamente associati.
5. Assorbimento di composti :
Hirsch et al., Hanno scoperto che quando lo zucchero di ferro viene somministrato ad un animale, il ferro viene assorbito dal complesso di Golgi (Kedrowsky). Van Teel ha dimostrato che i sistemi di Golgi assorbono anche composti di rame e oro. Kedrowsky ha dimostrato che il complesso di Golgi di Opalina può assorbire il bismutosio (composto di albumina e bismuto) e il protargol (composto di albumina e argento). Pertanto, Kirkman e Severinghaus affermano che il complesso di Golgi agisce come una membrana di condensazione per la concentrazione di prodotti in goccioline o granuli.
6. Formazione della parete cellulare vegetale :
La parete cellulare delle piante è costituita da fibrille che contengono prevalentemente polisaccaridi, insieme ad alcuni lipidi e proteine. Durante la citochinesi si forma una piastra cellulare tra i due nuclei figlie, e ha intorno a sé una membrana che in seguito diventa la membrana plasmatica delle cellule figlie. Esistono prove evidenti del fatto che i polisaccaridi si formano nel complesso del Golgi e trasferiti nella nuova parete cellulare che viene depositata mentre le cellule sono ancora in crescita.

Sostanze come le pectine e le emicellulose, che formano la matrice della piastra cellulare che separa le membrane plasmatiche, sono anche fornite dal complesso di Golgi.
7. Formazione di cristalli intracellulari :
Nell'isopodo marino, il Limnoria lingmorum, che è una forma scavatrice, sono presenti le midglands le cui cellule sono costituite da cristalli. Questi vanno fino a 30 A ° di lunghezza e 15 A ° di spessore. È stato dimostrato che questi cristalli sono formati dal complesso di Golgi e sono noti per contenere proteine e ferro. Sono privi di membrana e di solito di forma sferoidale. Sono interessati all'attività secretoria.
8. Formazione di goccioline di proteine del latte :
Nella ghiandola mammaria che allatta i topi vengono prodotte goccioline proteiche correlate al complesso di Golgi. Queste goccioline di solito si aprono sulla superficie cellulare mediante la fusione della membrana che le racchiude con la membrana plasmatica.
9. Formazione di lisosomi e vacuoli :
I lisosomi primari sono formati dalle membrane del Golgi allo stesso modo delle vescicole secretorie. Esistono buone prove del fatto che i dictyosomes accumulano enzimi idrolitici nelle loro regioni più mature. Alcuni vacuoli nelle cellule vegetali hanno dimostrato di contenere piccole quantità di enzimi idrolitici e si presume che siano derivati dal complesso di Golgi.
10. Formazione del pigmento:
In molte cellule tumorali e tumorali di mammiferi il complesso di Golgi è stato descritto come il sito di origine dei granuli di pigmento (melanina).
11. Regolazione del bilancio idrico :
È stata suggerita un'omologia tra il complesso del Golgi e il vacuolo contrattile del Metazoa inferiore e del Protozoo. Il vacuolo contrattile espelle l'acqua in eccesso dalla cella. In alcuni protozoi il complesso del Golgi si occupa anche della regolazione del bilancio idrico.
Origine del complesso di Golgi:
Sono state proposte tre diverse fonti da cui potrebbe emergere il nuovo complesso di Golgi:
1. Dal reticolo endoplasmatico :
Essner e Novikоff (1962) e Beams and Kessel (1968) hanno proposto che le cisterne del Golgi nascano dal pronto soccorso. Il reticolo endoplasmatico ruvido dopo aver sintetizzato proteine specifiche perde i ribosomi e cambia in ER liscio. Piccole vescicole transitorie si staccano da ER liscio. Questi migrano verso il dictyosome.
Raggiungendo la faccia formante di dictyosome questi si fondono per formare nuove cisterne e quindi contribuire alla sua crescita. Con la fusione di queste vescicole si formano continuamente nuove cisterne sulla faccia formante e sul viso in via di maturazione le vecchie cisterne si rompono in vescicole secretorie. Quindi il Golgi esibisce un fenomeno di flusso membranoso '.
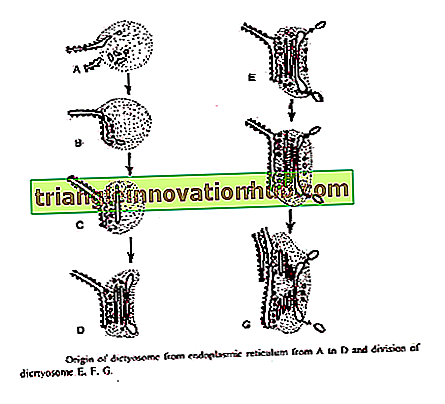

Bouch (1965) descrisse l'origine di Golgi dalla membrana esterna dell'involucro nucleare in alghe brune. I vespi sono staccati dalla membrana nucleare esterna che si fonde per formare cisterne sulla faccia formante del dictyosome.
Presenza di zone di esclusione in relazione a ER liscio o membrana nucleare, il verificarsi di zone di esclusione in semi dormienti di piante superiori e la formazione di dictosomi da queste zone in semi germinanti forniscono la prova a sostegno delle due precedenti teorie sull'origine di dictyosome.
3. Dalla divisione del dictyosome preesistente :
È stato osservato che durante la divisione cellulare in entrambe le piante e gli animali, il numero di dictiosomi aumenta e il numero di dictosomi in ogni cellula figlia appena dopo la divisione è quasi uguale al numero nella cellula madre prima della divisione, da lattine e altri osservazioni sulle cellule in divisione è stato presunto che anche i dicosomi si dividono durante la divisione cellulare.